연일층군의 두호층에서 산출된 서나무속(Carpinus)의 화석 포(苞)와 그 의미

Copyright © 2017 The Geological Society of Korea
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/ which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
초록
두호층에서 새로이 산출된 서나무속의 화석 포를 연구하고 다음과 같이 5종으로 분류했다; Carpinus subcordata Nathorst, C. stenophylla Nathorst, C. kodairae-bracteata Huzioka, C. miofargesiana Tanai and Onoe, and C. oblongibracteata Hu and Chaney. 뒤의 3종은 연일층군에서 처음 기록되었다. 한반도의 7개 신생대 신제3기 식물군 중에서 연일식물군의 서나무속의 종다양성이 가장 높게 나타난 것은 중기 마이오세에 포항 지역의 연평균 기온이 현재보다 약 2℃ 낮았던 것과 관련이 있을 것으로 판단된다. 이러한 환경 요인이 서나무속의 종다양성을 높이는데 중요한 역할을 했을 것으로 추정된다. 한반도의 전기-중기 마이오세 동안에 화석종의 수가 현생종에 비해 매우 많은 이유도 당시의 한반도의 연평균 기온이 전체적으로 현재보다 약간 낮았던 것과 관련이 있을 것으로 추정된다. 동아시아에서 서나무속의 종분화는 지역과 시대에 따라서 다르게 일어난 것으로 해석되었다. 일본에서는 올리고세 말기에 종분화가 왕성하게 일어났고 전기 마이오세에 들어와 크게 번성하였다. 반면에 한반도와 중국은 전기 마이오세에 종분화가 일어났고 중기 마이오세에 들어와 크게 번성한 것으로 추정된다. 한반도에서 Distegocarpus절의 C. cordata 형태군에 속하는 4종은 신제3기 동안에 모두 멸종하였다. C. subcordata Nathorst와 유연관계가 있는 까치박달(C. cordata)은 현재 한반도에 서식하고 있다. Eucarpinus절의 C. orientalis 형태군에 속하는 C. subyedoensis Konno는 신제3기 동안에 멸종하였다. 그러나 C. subyedoensis에 유연관계가 있는 개서나무(C. tschonoskii)는 현재 한반도에 서식하고 있다.
Abstract
This paper studied the fossil involucres of Carpinus from the Duho Formation of Yeonil Group, Korea and the involucres are classified into five species as follows; Carpinus subcordata Nathorst, C. stenophylla Nathorst, C. kodairae-bracteata Huzioka, C. miofargesiana Tanai and Onoe, and C. oblongibracteata Hu and Chaney. The occurrence of the last three species is their first record in the Yeonil Group. Compared to the seven Neogene floras of the Korean Peninsula, Carpinus in the Yeonil flora shows the highest species diversity. This probably reflects that the annual mean temperature of the Pohang area was about 2℃ lower than that of the present day. During the Early to Middle Miocene of the Korean Peninsula, the number of fossil species is much larger than that of modern species, which is also probably due to the lower annual mean temperature of the Korean Peninsula at that time. Speciation of Carpinus in the East Asia shows different patterns depending on area and period. In Japan, the speciation mostly took place at the end of the Oligocene, and Carpinus greatly flourished in the Early Miocene. On the other hand, it is presumed that the speciation in the Korean Peninsula and China took place mostly in the Early Miocene and Carpinus greatly flourished in the Middle Miocene. Four species belonging to the form group C. cordata of section Distegocarpus on the Korean Peninsula were all extinct during the Neogene. C. cordata, which has a relationship with C. subcordata Nathorst, is present in the Korean Peninsula. C. subyedoensis Konno belonging to the form group C. orientalis of section Eucarpinus was extinct during the Neogene. However, C. tschonoskii, which has a relationship with C. subyedoensis, is present in the Korean Peninsula.
Keywords:
Carpinus, fossil involucre, Yeonil Group, Miocene, Duho Formation, Distegocarpus, Eucarpinus키워드:
서나무속, 화석 포, 연일층군, 마이오세, 두호층, 디스테고카르푸스, 유카르피누스1. 서 론
자작나무과(Betulaceae)는 온대 지역에서 가장 우세한 그룹이고, 자작나무속(Betula), 서나무속(Carpinus), 오리나무속(Alnus), 개암나무속(Corylus), 새우나무속(Ostrya) 등이 가장 대표적이다(Tanai, 1961). 그중에서 서나무속(Carpinus)은 북반구의 온대와 아열대지역에 25종(Worobiec and Szynkiewicz, 2007) 내지 35종(Jones and Luchsinger, 1986; Ko, 1991)이 분포하며 후자의 대부분은 중국에 분포한다(Hiller and Coombes, 2002). 이들은 유럽에서 동아시아까지, 히말라야의 남부까지 그리고 북아메리카에서 중앙아메리카에 걸쳐 분포하며(Chang and Jeon, 2004), 아열대 지역에 속하는 인도와 중앙아메리카의 산간 지역에도 분포한다(Tanai, 1961, 1972; Stults et al., 2002). 한반도에도 5종 (Lee and Lee, 1991; Lee, 2006) 내지 4속 2변종(Jeon and Chang, 1997)이 서식하고 있을 정도로 잘 알려져 있는 식물이다. 자작나무과 식물의 선조형은 백악기 후기에 이미 출현한 것으로 알려져 있지만(Flora of North America, 1997; Chenet al., 1999), 서나무속의 선조형은 그보다 조금 늦은 신생대 고제 3기에 나타났고(Pigg et al., 2003), 그 최초의 화석 기록은 아시아와 유럽의 에오세 지층에서 보고되었다(Endo, 1950; Tanai, 1972; Uemura and Tanai, 1993; Wilde and Frankenhäuser, 1998). 세계적으로 고제3기의 지층에서 많이 산출되고 있다.
한반도에서 기록된 신제3기의 서로 다른 지역의 7개 식물화석군(Huzioka, 1972)은 모두 서나무속의 화석종을 다수 포함하고 있다. 초기의 화석 기록은 Kryshtofovich (1921)가 일본 동북대학교에 소장된 장기식물군의 화석을 관찰하고 보고한 10종을 비롯하여 Tateiwa (1924a, 1924b)가 남한에 분포하는 장기층군에서, Kanehara (1935, 1936)가 연일층군에서, 그리고 Endo(1938a, 1938b)가 북한 지역에 분포하는 명천층군과 고건원 탄전에서 보고한 화석 목록에서 찾아볼 수 있다. 이어서 Endo (1939, 1950), Huzioka (1943, 1954), Tanai (1952), Chun (1982), Ablaev et al. (1993) 및 Lim et al. (1994)의 연구에서도 서나무속 화석종의 기록이 보편적으로 나타난다. Huzioka (1972)는 기존에 한반도에서 기록된 서나무속의 화석종을 모두 재검토하여 총 8종으로 정리하였다. Huzioka (1972)의 종합적 연구 이후, Ablaev et al. (1993)은 북한의 고건원 지역에서 2종을, 그리고 Lim et al. (1994)은 북한에 분포하는 제3기의 새로운 화석 산지로부터 7종의 화석종을 각각 보고하였다. 현재까지 연일층군에서 기재된 서나무속의 화석종은 Carpinus hokoensis Endo, C. stenophylla Nathorst, C. subcordata Nathorst 및 C. sp.가 있다(Kanehara, 1936; Endo, 1950; Tanai, 1952; Huzioka, 1972; Chun, 1982). 이들은 잎 또는 포에 근거하여 기재되었다. 포는 모양과 엽맥 등이 종에 따라 특징이 있기 때문에 종의 식별에 자주 이용되고, 현생종과의 유연관계를 조사하는 데도 유효하기 때문에 매우 중요한 화석이다(Tanai, 1972).
한편, 한반도에서 서나무속의 화석종은 잎이나 포이외에도 화분과 목재에 대한 연구도 상당히 진행되었다. 연일층군의 화석 화분에 대한 연구는 Takahashi and Kim (1979), Chang et al. (1988), Yamanoi (1992) 및 Chung and Choi (1993) 등에 의해 이루어졌다. 이들의 연구에 의해 연일층군의 모든 층에 서나무속의 화분이 다수 포함되어 있다는 것이 확인되었다. 화석 목재는 현재까지 연일층군에서 산출된 적은 없지만, 시대적으로 더 오래된 장기층군의 하부함탄층에서 C. donghaeensis Jeong and Kim과 C. sp.가 보고된 바 있다(Kim et al., 2008; Jeong et al., 2009).
최근에 포항시 북부항 부근에 분포하는 두호층의한 화석 산지에서 형태가 다른 여러 종류의 서나무 속의 화석 포가 새로이 발견되었다. 이들 화석포에 대한 연구는 서나무속의 종다양성을 비롯하여 현생종과 화석종의 유연관계에 대한 정보, 이웃한 중국과 일본의 비슷한 시대의 지층에서 기록된 화석종과의 비교를 통한 고식물지리구의 추정, 서식환경과 분포를 통한 고기후의 추정 등에 중요한 기초 자료를 제공하였다. 따라서 이 연구는 새로이 발견된 포의 형태적 특징을 기재하고 그 의미에 대해서 논의하고자 한다.
2. 재료 및 연구 방법
Carpinus속의 현생종은 Jeon and Chang (2000)이 언급한 바와 같이, 2개의 절(Distegocarpus와 Eucarpinus), 또는 2개의 아속(Distegocarpus와 Eucarpinus)과 2개의 절(Elongatae와 Brachyspicae), 또는 2개의 절(Distegocarpus와 Carpinus)과 3개의 아절(Carpinus, Monbeigianae, Polyneurae) 등의 여러 가지 방법으로 세분되고 있지만, 화석종의 경우는 일반적으로 절이나 아속명까지 세분하지 않는다. 따라서 여기에서는 속명과 종명으로만 구분한다. Carpinus속의 한국명은 서나무속(Lee and Lee, 1991; Lee, 2006), 서어나무속(Ko, 1991) 또는 서나무속/서어나무속(Yun, 2017) 두 개를 병용하여 표기하고 있는데, 여기에서는 서나무속의 이름을 사용한다. 그리고 과실에 대한 명칭도 소견과(Lee, 2006), 또는 견과(Ko, 1991; Lee and Lee, 1991)가 있지만 후자를 따라서 기재했다. 그리고 포에 대한 명칭은 총포(總苞; Lee and Lee, 1991) 또는 포(苞; Lee, 2006; Yun, 2017)가 있지만 후자를 따랐다.
연일층군에서 채집된 서나무속의 포 표본은 총10개이다. 이들은 모두 인상으로 보존되었지만 보존상태가 양호한 편이다. 포의 경우 엽맥과 외부 형태의 특징만이 종을 식별할 수 있는 유일한 기준이다. 따라서 이 연구에서는 포의 형태, 엽맥, 거치의 유무와 모양, 크기, 견과의 크기 등을 계측하여 정리하고 문헌적 연구를 통해서 종 분류를 하였다. 이 연구에 사용된 표본은 모두 공주대학교(KNU) 표본실에 보관되어있다.
3. 선행 연구
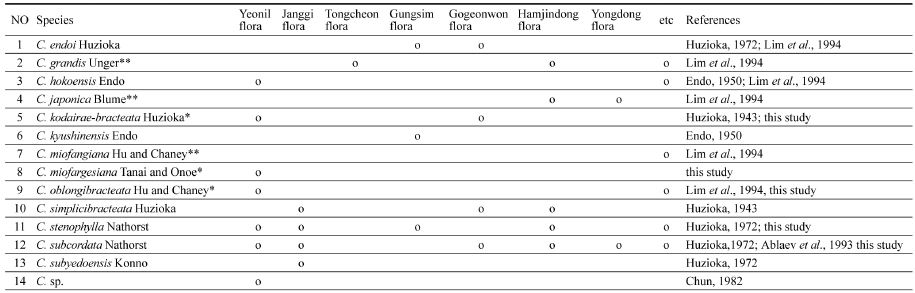
한반도에 분포하는 신제3기층에서 2016년까지 서나무속의 화석 잎 또는 포에 근거하여 기재된 종은 표 1과 같이 12종(1개 미지정종 제외)이다.

Fossil species of Carpinus described by previous works from the Neogene strata of the Korean Peninsula.
4. 연구 지역의 지질과 시대
Tateiwa (1924b)는 처음 연일층군의 지질과 층서를 조사하였다. 그 후 국내의 여러 연구자들에 의해 수차례 층서적 연구가 수행되었으며(Um et al., 1964; Yoon, 1975; Yun, 1986; Kim, 1987), 층서 구분과 층명은 Lee and Lee (1986)의 층서대비표에서 보는 바와 같이 조금씩 서로 다르다. 이 연구에서는 Um et al. (1964)의 층서 분류를 따랐다. Um et al. (1964)에 의하면, 연일층군은 6개 층으로 세분되어 있지만, 화석 포가 산출된 북부항 부근에는 3개 층이 분포한다. 그림 1은 연구 지역의 지질도와 화석 산지를 나타낸 것이다. 서나무속의 포는 모두 두호층에서 산출되었다. 두호층은 이암, 셰일 및 사암의 호층으로 구성되며 암석의 색은 백갈색이며 층의 두께는 150~200 m에 이른다(Um et al., 1964). 연일층군은 내만성 내지 연안성 천해 환경에서 형성된 것으로 추정되며(Huzioka, 1972), 해서 척추 및 무척추동물, 미화석 및 대형식물이 화석으로 많이 산출되고 있다. 그 상세한 내용과 출전은 Lee (1987)와 Kim (1996)에 의해 정리되었다.

Geological map of the Pohang Basin (Um et al., 1964, recited in Kim et al., 2017) and fossil locality.
연일층군의 지질 시대는 연체동물, 규조, 유공충, 화분을 비롯한 대형 식물화석에 의하면 대체로 중기 마이오세에서 후기 마이오세로 보고 있지만(Bong, 1981; Yoon, 1982; Choi et al., 1984; Yun, 1986; Yamanoi, 1992; Chung and Choi, 1993), 후기 올리고세 내지 전기 마이오세의 의견도 있다(Chun, 1982). 한편, 유공충 및 유기질 미화석의 연구에 의한 전기에서 중기 마이오세(Kim, 1990; Yun et al., 1991) 또는 규조를 비롯한 미화석에 의한 중기 마이오세(Lee et al., 1991) 등의 견해도 있다. 화산암의 절대연대 측정에 의한 연일층군의 연대는 15 Ma로 보고되기도 했다(Lee et al., 1992).
5. 연구 결과
10개의 화석 포에 대한 연구 결과 다음과 같이 5종으로 분류되었다. 그중에서 2종은 기존에 알려진 종이고, *표가 붙어 있는 3종은 연일층군에서 처음 산출되었다.
Carpinus kodairae-bracteata Huzioka*
Carpinus miofargesiana Tanai and Onoe*
Carpinus oblongibracteata Hu and Chaney*
Carpinus stenophylla Nathorst
Carpinus subcordata Nathorst
Systematic description
Betulaceae Gray
Carpinus Linnaeus 1737
Carpinus kodairae-bracteataHuzioka 1943
Fig. 2a
1943 Carpinus kodairae-bracteata Huzioka, p. 290, pl. 14, figs. 14, 15, 15a
1954 Carpinus kodairae-bracteata Huzioka, p. 119
1961 Carpinus kodairae-bracteata Huzioka, Tanai, p. 293, pl. 11, fig. 4
1964 Carpinus kodairae-bracteata Huzioka, p. 74, pl. 7, fig. 1
1972 Carpinus kodairae-bracteata Huzioka, p. 47, pl. 4, fig. 11
Material: KNU-2016-0001
Description: Involucre is palmately tri-lobed. The median lobe is the largest, lanceolate in shape, 1.8 cm long and 1.1 cm wide, acute at apex, coarsely serrate at lateral margin; lateral two lobes are small, trigonal in shape, unequal in size, and slightly serrate at margin. Primary veins are about 6 in number, extending from the base, then entering into the apex of lobes or marginal teeth, the median primary is the longest, a few distinct secondaries are diverging from the median primary at acute angles, ending in the margin teeth or margin, tertiaries are not preserved. Base is asymmetrically rounded. Stalk is short, about 3.2 cm long and 1.4 cm wide.
Occurrence and distribution: This species occurs in the Duho Formation of the Yeonil Group. This species has also been known in the Neogene floras of Japan (Tanai, 1964).
Remarks: The most distinctive characters of the present specimen are the tri-lobed and asymmetrical involucre with an acute apex and six primary veins. These characters are most similar to those of Carpinus kodairae-bracteata established by Huzioka (1943) from the Gogeonwon Coalfield of North Korea, but the latter slightly differs from the present specimen in that they have numerous primary veins (about 10 in number) and acutely pointed apex. Although the number of primary veins in the presnt specimen is fewer than that of original specimens, we regarded it as Carpinus kodairae-bracteata Huzioka.
C. kodairae-bracteata Huzioka somewhat resembles those of C. miocenica Tanai described by Tanai and Onoe (1961) from the Mio-Pliocene floras of Japan in its tri-lobed shape, but the former is distinguished from the latter by the overall shape and venation pattern of the lobe.
As mentioned by Tanai (1961) and Huzioka (1972), the involucre of C. kodairae-bracteata Huzioka is mostly similar to the those of modern C. laxiflora Blume of Korea, Japan, and mainland China.
Carpinus miofargesiana Tanai and Onoe 1959
Fig. 2b
1955 Carpinus miofargesiana Tanai, pl. 5, figs. 15-17
1959 Carpinus miofargesiana Tanai and Onoe, p. 278, pl. 5, figs. 4-6, 9-11
Material: KNU-2016-0002
Description: Involucre is obliquely ovate in shape, asymmetrically rounded at the base, 1.5 cm long and 0.8 cm wide, acute at apex. Margins are lobately serrate on the larger half, nearly entire in the lower part and slightly serrate in the upper part of the smaller half. Primary veins are about 5 in number, extending from the base, then entering into the apex of lobes or marginal teeth, the median primary is the longest; a few distinct secondaries diverge from the median primary at wide angles and enter into smaller teeth or connect with adjacent primary veins; tertiaries are not distinct, but a few form a transverse network. Stalk is short, about 3.2 cm long and 1.4 cm wide, 1.5 cm long and 1.8 cm wide. Nutlet is poorly preserved, ovate in shape, 2.5 mm long and 1.8 mm wide.
Occurrence and distribution: This species is only known in the Duho Formation of the Yeonil Group.
Remarks: Carpinus miofargesiana was established based on three forms of involucre by Tanai and Onoe (1959, plate V, figs. 9-11) from the Miocene flora distributed in the Joban Coalfield of Japan. The present specimen is most similar in shape to the specimen of fig. 10, but the number primary veins varies from specimen to specimen (Tanai, 1961). The Korean specimen also shows some variation in the number of primaries, but is assigned to C. miofargesiana Tanai and Onoe. The leaves of C. miofargesiana Tanai and Onoe have been known in the Buhwa and Saebyeol Series of North Korea (Lim et al., 1994).
Carpinus oblongibracteata Hu and Chaney 1940
Fig. 2e-j
1940 Carpinus oblongibracteata Hu and Chaney, p. 33, pl. 12, figs. 7, 8
Material: KNU-2016-0006, 0007, 0008, 0009, 0015, 0016
Description: Six specimens of involucre were obtained. Involucre is asymmetrically ovate in shape, 1.7-1.9 cm long and 0.7-0.8 cm wide, acute at apex. Margin is lobately serrate or slightly serrate on the larger half, slightly serrate on the smaller half. Primary veins range from 5 to 7 in number, extending from the base, then entering into apex of lobes or teeth; the median primary is the longest, a few distinct secondaries diverge from the median primary at wide angles and enter into smaller teeth; tertiaries are not preserved. Base is rounded. Stalk is short, about 1.4-2.5 mm long and 09-1.4 mm wide. Nutlet is poorly preserved, ovate in shape, 2.7-3.2 mm long and 1.8-2.3 mm wide.
Occurrence and distribution: This species is known in the Duho Formation of the Yeonil Group of South Korea (Huzioka, 1972) and Buhwa Series of Hamgyeongbug-do, North Korea (Lim et al., 1994).
Remarks: The present specimens agree well with those of Carpinus oblongibracteata established by Hu and Chaney (1940) from the Miocene flora of Shantung, China. The involucres of C. oblongibracteata are similar in shape to those of modern C. tschonoskii Maxmoxicz distributed in Japan and China (Hu and Chaney, 1940; Inst. Bot. and Inst. Geol. & palaeont., 1978). The latter species is also distributed in the southern part of the Korean Peninsula (Yun, 2017).
Carpinus stenophylla Nathorst 1883
Fig. 2d
1883 Carpinus stenophylla Nathorst, p. 41, pl. 3, fig. 16
1939 Carpinus japonica Blume, Endo, p. 340, pl. 23, figs. 3, 4
1950 Carpinus protojaponica Nathorst, Endo, p. 340, pl. 23, figs. 3, 4
1954 Carpinus carpinoides Makono, Huzioka, p. 119
1994 Carpinus protojaponica Nathorst, Lim et al., p. 138, pl. 68, fig. 9
Material: KNU-2016-0010
Description: This species was represented by a single specimen. Involucre is asymmetrically ovate to oblong in shape, unequally rounded at the base, 1.8 cm long and 0.8 cm wide. Apex is not preserved. Margin is nearly entire and serrate on the larger half in a few specimens. Primary veins are poorly preserved, at least 5 veins extend from the base, then enter into apex or teeth, median primary is the longest, and secondaries are not preserved. Stalk is stout, 3.6 mm long and 1.8 cm wide. Nutlet is large, ovate in shape, 3.6 mm long and 3.2 mm wide.
Occurrence and distribution: This species is known in the Duho Formation of the Yeonil Group and also commonly known in the Miocene floras of the Korean Peninsula (Huzioka, 1972; Ablaev et al., 1993).
Remarks: The present specimen is characterized by its a few serration and ovate-oblong shape. These characters agree well with those of C. stenophylla Nathorst described from the Miocene floras of the Korean Peninsula (Huzioka, 1972). The similar fossil involucres have been described from Neogene floras in Japan and Korea and given various names as seen in the synonym list. This species is mostly similar to the extant C. japonica Blume of Japan (Huzioka, 1972).
Similar fossil involucres referred to Protojaponica Nathorst were reported from the Saebyeol and Gocham Series of North Korea (Lim et al., 1994), and judging from the size and shape of involucre, they may be assinable to C. stenophylla Nathorst.
Carpinus subcordata Nathorst 1883
Fig. 2c
1883 Carpinus subcordata Nathorst, p. 39, pl. 2, figs. 13-18, 20; Huzioka, 1972, pl. 4, figs. 1, 1a
1943b Carpinus erosa Bl. ellipticibracteata Huzioka, p. 290, pl. 19, figs. 6, 7, 7a, 8, 8a, 9, 9a; Huzioka, 1954, p. 119
1954 Carpinus miocordata Hu and Chaney, p. 31, pl. 12, figs. 1, 2, 11; Huzioka, 1954, p. 120, pl. 13, figs. 5, 6
Material: KNU-2016-0011
Description: Involucre is ovate to elliptical in shape, 2.3 cm long and 1.1 cm wide, bluntly pointed at the apex. Margins are coarsely serrate in the middle and upper part. Primary veins are 4 in number, extend from the base, then enter into apex or teeth; median primary is the longest; secondaries diverge from the median primary at wide angles and enter into smaller teeth; tertiaries are not preserved. Base is symmetrically rounded. Stalk is short, 1.8 mm long and 1.8 mm wide. Nutlet is not preserved.
Occurrence and distribution: This species is known in the Duho Formation of the Yeonil Group and also commonly known in the Miocene floras of the Korean Peninsula (Huzioka, 1972).
Remarks: The fossil leaves and involucres of C. subcordata Nathorst have been described in Neogene and Pleistocene floras in Japan and Korea (Tanai, 1961; Huzioka, 1972) and given various names as seen in the synonym list. The present specimen agrees well with those of C. subcordata Nathorst described by Huzioka (1972). C. subcordata Nathorst is also one of the most common species in the Neogene floras of northern Asia (Tanai, 1961; Huzioka, 1972; Sun et al., 2003).
As mentioned by Huzioka (1972), the involucre of C. subcordata Nathorst resembles those of extant C. cordata Blume which are widely distributed in Korea, Japan, and China. Sun et al. (2003) firstly described cuticular structure of C. subcordata Nathorst from the Miocene strata of Yunnan Province, China. According to them, cuticular characteristics of C. subcordata Nathorst is very similar to the extant C. cordata Blume. Similar fossil involucres assigned to C. cordata Blume were reported from the Saebyeol and Gocham Series of North Korea (Lim et al., 1994), and judging from the size and shape of involucre, they should be included in C. subcordata Nathorst.
The involucre of C. subcordatacarpinus kitamiensis established by Uemura and Tanai (1993) from the Oligocene Wakamatsuzawa Formation of Hokkaido, Japan, is somewhat similar in shape to the present species, but the former differs from the latter by its wide elliptic involucres with a rounded apex and base.
6. 토 의
연일식물군에서 기록된 서나무속의 화석종은 3종(1개의 미결정종 미포함)이었지만, 이번의 연구로 Carpinus kodairae-bracteata Huzioka, C. miofargesiana Tanai and Onoe, C. oblongibracteata Hu and Chaney의 3종이 추가되어 총 6종에 이른다. 이들을 포함한 한반도의 신제3기층에서 기록된 화석종은 표 2와 같이 총 13종(1개의 미결정종 미포함)이다. 그중에 3종은 잎에 의해 나머지 10종은 포에 의해 각각 기재되었다. 연일층군에서 기재된 서나무속의 화석종의 수는 신제3기의 7개 식물군(Huzioka, 1972) 중에서 가장 많다. 이것은 여러 가지 환경 요인 중에서 연평균 기온과 가장 밀접한 관계가 있을 것으로 생각된다. 포항 지역은 신제3기의 다른 식물군들보다 위도적으로 가장 남쪽에 위치하여 상대적으로 가장 따뜻한 지역이며 최근에 측정한 포항 지역의 연평균 기온은 평균 14.4℃이다(Lee et al., 2003). 같은 지역에서 연일식물군이 번성할 당시의 연평균 기온은 Huzioka(1972)의 대형화석 식물의 전연률(37%)과 Wolfe (1978)의 전연률과 연평균 기온과의 상관관계(기온 = 전연률 × 0.306 + 1.14)를 이용하면 구할 수 있다. 이 방법을 이용하여 구한 연평균 기온은 대략 12.4℃이며, 이 값은 현재보다 약 2℃정도 낮다. 이러한 환경은 서나무속의 식물이 번성하는데 현재보다 더 적합한 환경이었으며, 이것 때문에 종다양성이 높게 나타난 것으로 판단된다. 한편, Chung and Choi (1993)는 위에 언급한 것과 똑같은 방법과 같은 자료를 이용하여 연평균 기온을 구하고 15℃로 추정했다. 그러나 Kim and Choi (2008)가 이미 언급한 바와 같이, 연평균 기온이 15℃가 되려면 전연률이 최소 45%를 넘어야 가능하다. 이와 같이 높게 나타난 이유는 아마도 Wolfe (1978)의 그래프에서 연평균 기온을 잘못 읽었을 가능성이 크다. 또한, 연일식물군의 하부에 놓이는 장기식물군의 전연률은 약 31%이고(Huzioka, 1972), 이 값에 대응하는 연평균 기온은 대략 10.5℃이다. 이것은 연일식물군보다 약 2℃정도 낮다. 이것은 전기 마이오세의 장기식물군에서 중기 마이오세의 연일식물군 쪽으로 가면서 연평균 기온이 점차로 상승했다는 것을 의미한다. 이러한 기온 상승은 장기식물군에 상록수가 한 종도 없는 것에 비해 연일식물군에는 10종의 상록수가 포함되어 있다는 것과 일치한다(Huzioka, 1972). Wolfe (1978)의 제3기의 연평균 기온의 변화곡선을 보면, 북아메리카 지역에서 팔레오세에서 올리고세 동안에 연평균 기온은 급격하게 상승과 하강을 반복하면서 점차로 하강하였고, 올리고세 이후부터 제4기까지는 완만하게 상승과 하강을 반복하면서 점차적으로 하강한 것으로 나타난다. 이와 비슷한 기온변화 패턴이 동아시아 지역에서도 알려져 있다(Tanai, 1984, 1991). 특히, 마이오세 전기부터 중기에는 기온이 점차로 상승하다가 중기 이후에는 다시 하강한다. 장기식물군과 연일식물군이 번성한 시기는 마이오세의 기온의 상승기에 해당한다. 그렇지만 전기-중기 마이오세 동안에 한반도의 연평균 기온은 전체적으로 현재보다 약간 낮았다고 본다.
Fossil species of Carpinus described from the Neogene strata of the Korean Peninsula based on the involucres(*; new occurrence in this study, **; leaf only).

Fossil involucres of Carpinus discovered from the Miocene Duho Formation. a; C. kodairae-bracteata Huzioka (KNU-2016-0001), b; C. miofargesiana Tanai and Onoe (KNU-2016-0002), c; C. subcordata Nathorst (KNU-2016-0011), d; C. stenophylla Nathorst (KNU-2016-0010), e-j; C. oblongibracteata Hu and Chaney (KNU-2016-0006, 0007, 0008, 0009, 0015, 0016).
한편, 7개 식물군에서 기록된 13종의 화석종에 동종이명이 포함되어 있을 가능성을 감안하더라도 현재 한반도 전역에 분포하는 5종의 현생종에 비하면(Lee, 2006) 화석종의 수가 매우 많다. 이것은 위에서 언급한 바와 같이 전기-중기 마이오세 동안에 장기식물군과 연일식물군이 번성할 당시 한반도의 연평균 기온은 전체적으로 현재보다 약간 낮았고, 이러한 환경이 서나무속의 식물이 번성하는데 현재보다 더 적합한 환경이었기 때문이라고 추정된다.
표 2와 같이 포항 지역의 연일식물군에서는 6종이 알려져 있지만, 시대적으로 더 오래된 장기식물군에서는 4종이 알려져 있다. 이와는 반대로 장기 식물군에 대비되는 일본의 전기 마이오세의 Aniai형 식물군은 11종의 화석종을 포함하며, 연일식물군에 대비되는 일본의 중기 마이오세의 Daijima형 식물군은 6종의 화석종을 포함한다(Tanai, 1961). 그리고 중국의 중기 마이오세 식물군에서는 10종이 알려져 있다(Inst. Bot. and Inst. Geol. & Palaeont. Acted. Sin, 1978). 이는 동아시아에서 서나무속의 종분화가 지역과 시대에 따라 다르게 일어났다는 것을 보여준다. 즉, 일본에서는 올리고세 말기에 종분화가 왕성하게 일어났고 전기 마이오세에 들어와 크게 번성했지만 중기에 이르러 이들의 대부분은 사라졌다. 그리고 한반도와 중국은 전기 마이오세 말기에 종분화가 활발하게 일어나고 중기에 번성했지만, 그들의 대부분은 마이오세 말기에 멸종된 것으로 보인다. 이러한 경향은 Tanai (1972)의 연구에서도 비슷하게 나타난다.
Berger (1953)는 세계 각지에 서식하는 서나무속의 현생종과 화석종을 조사하고 이들 간의 계통관계 및 분포와 변천을 명백히 밝혔다. 그는 현생종과 화석종을 포의 특징에 의해서 8개 형태군(form group)으로 나누고 이들을 서나무속 내의 2개의 절(section), 즉 Distegocarpus (Siebold et Zuch.) Sargent와 Eucarpinus Sargent에 포함시켰다. 전자는 C. cordata, C. japonica 및 C. rankanensis 형태군을 포함하고, 후자는 C. betulus, C. caroliniana, C. macrocarpa, C. orientalis 및 C. tschonoskii 형태군을 포함한다. Distegocarpus에 속하는 그룹은 포가 대체로 넓은 난형에서 타원형이며 거치가 상부 엽연에 나타나며 하부에서는 포가 열편으로 갈라지지 않는다. 반면에 Eucarpinus에 속하는 그룹은 포의 형태가 넓은 난형이고 일부는 하부에서 뚜렷하게 3개의 열편으로 갈라진다(Berger, 1953). Tanai (1972)는 Berger (1953)의 자료에 일본에서 얻어진 자료를 추가하여 제3기 동안에 일본에서 서나무속의 시대적 변천과 현생종간의 유연관계 등을 상세히 나타냈다.
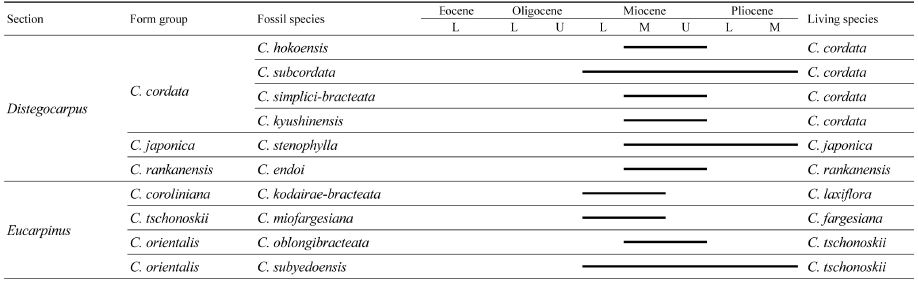
표 3은 Tanai (1972)와 비슷한 방법을 사용해서 한반도의 신제3기 식물군에서 기록된 13종 중 화석 포에 의해 기재된 10종에 대해 시대적 분포 및 종간 유연관계를 나타낸 것이다. Distegocarpus절에는 C. cordata 형태군, C. japonica 형태군 및 C. rankanensis 형태군에 6종이 포함되어 있다. C. cordata 형태군에 속하는 4종 중에서 C. subcordata Nathorst를 제외한 3종은 모두 후기 마이오세에 멸종되었다. C. subcordata Nathorst는 한반도를 포함한 일본과 중국의 마이오세에서 플라이오세까지 공통적으로 산출되며, 현생의 까치박달(C. cordata Blume)과 유연관계가 있다(Tanai, 1972; Inst. Bot. and Inst. Geol. & Palaeont., Acad. Sin., 1978). 중국 윈난성(Yunnan)의 마이오세 지층에서 보고된 C. subcordata Nathorst의 큐티클의 구조가 현생의 C. cordata Blume (까치박달)의 그것과 매우 비슷하다는 것이 알려져 있다(Sun et al., 2003). 이것으로 보아 C. subcordata Nathorst가 진화과정을 거쳐 현생의 까치박달이 출현한 것으로 판단된다. 까치박달은 현재 한반도, 중국 및 일본에 걸쳐 분포하고 있다(Jeon and Chang, 2000). C. japonica 형태군에서 플라이오세까지 지속된 화석종은 C. stenophylla Nathorst 1종뿐이다. 이종은 현재 일본에만 분포하는 C. japonica Blume에 유연관계가 있다(Tanai, 1972). 이것에 대해 Jeon and Chang (2000)은 Tateiwa (1976)의 문헌을 인용하여 제3기부터 C. japonica Blume가 한반도에 서식한 것으로 보고 이것이 후에 한반도에서는 쇠퇴하고 일본에만 잔존한 것으로 해석했다. 그렇지만 한반도에서 기록된 C. japonica (Endo, 1939)와 C. protojaponica (Endo, 1950) 등은 후에 Huzioka (1972)에 의해 C. stenophylla로 종명이 모두 변경되었다. C. stenophylla는 플라이오세까지 나타나며, 일본과 한반도에서 공통적으로 산출된다. 플라이오세 이후 이종은 한반도에서 더 이상 나타나지 않는다. C. rankanensis 형태군에 속하는 C. endoi Huzioka는, Endo (1950)가 C. rankanensis Hayata로 보고한 종을 Huzioka (1972)가 신종으로 기재하면서 종명을 변경하였다. C. rankanensis Hayata는 현재 타이완에만 서식하고 한반도와 일본에는 서식하지 않는 종이다.
Stratigraphic distribution of fossil Carpinus species described from the Neogene floras based on the fossil involucres [data based on Berger(1963) and Tanai (1972)]
한편 Eucarpinus절에 속하는 화석종은 C. kodairaebracteata Huzioka, C. oblongibracteata Hu and Chaney 및 C. miofargesiana Tanai and Onoe 및 C. subyedoensis Konno의 4종이 있다. 그중에서 C. subyedoensis Konno를 제외한 나머지 3종은 모두 후기 마이오세 이후 산출되지 않는다. C. kodairae-bracteata는 한반도와 일본의 마이오세 지층에서 화석으로 산출되며 현재 한반도와 일본에 분포하는 서나무(C. laxiflora Blume)와 유연관계가 있다(Tanai, 1961). C. oblongibracteata Hu and Chaney는 현생의 개서나무(C. tschonoskii Maximowicz)에 가깝다(Beijing Inst. Bot. & Nanjing Inst. Geol. Paleonto., 1978). C. miofargesiana Tanai and Onoe는 현재 중국의 중부와 남부 지역에 분포하는 C. fargesiana Winkle과 유사하다(Tanai, 1972). 이것은 후기 마이오세 이후 이 종이 한반도에서는 멸종되었지만 중국에서는 진화과정을 거쳐 C. fargesiana Winkle이 출현한 것으로 볼 수 있다.
C. subyedoensis Konno는 한반도와 일본의 신제3기식물군에서 흔하게 나타나지만 중국의 신제3기 식물군에는 나타나지 않으며, 현생의 개서나무(C. tschonoskii Maximowicz)와 유연관계가 있다(Huzioka, 1972; Tanai, 1972), 따라서 C. subyedoensis Konno는 신제3기 동안에 진화하여 현생의 개서나무가 출현한 것으로 볼 수 있다. 현재, 개서나무는 중국, 한반도 및 일본까지 분포하는 것으로 보아, 플라이오세 이후 진화과정을 거치는 동안 서식지가 중국으로까지 확대되었다고 하는 Jeon and Chang (2000)의 의견과 일치한다.
7. 결 론
연일층군의 두호층에서 새로이 산출된 화석 포에 대해 연구하고 다음과 같이 5종으로 분류하였다; C. kodairae-bracteata Huzioka, C. miofargesiana Tanai and Onoe, C. oblongibracteata Hu and Chaney, C. stenophylla Nathorst 및 C. subcordata Nathorst. 뒤의 3종은 연일층군에서 처음 발견된 종이다. 연일층군에서 기록된 서나무속의 수는 한반도의 제3기의 서로 다른 지역의 식물화석군 중에서 종다양성이 가장 높다. 이것은 중기 마이오세에 포항 지역의 연평균기온이 현재보다 약 2℃ 정도 낮았던 것과 관련이 있을 것이며, 또한 이러한 기온은 서나무속의 다양성을 높이는데 중요한 역할을 했을 것으로 추정된다. 한반도에서 전기-중기 마이오세에 화석종의 수가 현생종의 수에 비해 매우 많은 이유는 당시의 한반도의 연평균 기온이 현재보다 약간 낮았고, 이러한 환경이 서나무속의 식물이 번성하는데 현재보다 더 적합한 환경이었기 때문으로 추정된다. 한반도에서 Distegocarpus절에 속하는 화석종은 대부분 멸종하였지만, 현생의 까치박달과 유연관계가 있는 C. subcordata Nathorst는 동아시아의 각 지역에서 진화과정을 거쳐 현재까지 잔존한 것으로 판단된다. C. stenophylla는 플라이 오세 이후 한반도에서 멸종되었다. Eucarpinus절의 화석종은 C. subyedoensis Konno를 제외한 나머지는 모두 후기 마이오세에 멸종했다. 이종은 신제3기 동안에 진화되어 현생의 개서나무가 출현한 것으로 판단된다. 동아시아에서 서나무속은, 일본에서는 올리고세 말기에 종분화가 왕성하게 일어났고 전기 마이오세에 들어와 크게 번성했지만 중기에 이르러 이들의 대부분은 사라진 것으로 해석된다. 반면에 한반도와 중국은 전기 마이오세에 종분화가 일어나 중기에 번성하고 그들의 대부분은 마이오세 말기에 멸종된 것으로 해석된다.
Acknowledgments
본 논문이 나오기까지 논문을 심사해주고 내용의 미비점에 대하여 상세한 지적과 건설적인 비평을 주신 전희영 박사님과 충북대학교 이동찬 교수님에게 깊은 감사를 드립니다.
References
- Ablaev, A.G., Sin, E.U., Vassiliev, I.G., and Lu, Z.M., (1993), Miocene of the North Korea and the South Primorye (beds with Engelhardia), FEB RAN, p1-140, (in Russian).
- Berger, W., (1953), Studien zur systematik und geschichte der gattung Carpinus, Botaniska Notiser, 1, p1-47.
- Bong, P.Y., (1981), Palynology of the Janggi area. Report of Geoscience, Mining Resources, 10, p7-16, (in Korean with English abstract).
-
Chang, C.-S., and Jeon, J.I., (2004), Foliar flavonoids of the most primitive group, sect. Distegocarpus within the genus Carpinus, Biochemical Systematics and Ecology, 32, p35-44.
[https://doi.org/10.1016/S0305-1978(03)00186-8]
- Chang, N.-K., Kim, K.-W., and Kim, J.-G., (1988), Analyses of fossil pollen of Tertiary Miocene in the Yeonil Area, Korea, Korean Journal Ecology, 11, p137-144.
-
Chen, Z.H., Manchester, S.R., and Sun, H.Y., (1999), Phylogeny and evolution of the Betulaceae as inferred from DNA sequences, morphology, and paleobotany, America Journal of Botany, 86, p1168-1181.
[https://doi.org/10.2307/2656981]
- Choi, D.K., Bong, P.Y., and Kim, B.K., (1984), Tasmanitids from Idong and Pohang (Miocene) Formations for Korea, Journal of the Geological Society of Korea, 20, p215-221.
- Chun, H.Y., (1982), Plant fossils from the Tertiary Pohang Sedimentary Basin, Korea, Korea Institute of Energy and Resources, 14, p7-23, (in Korean with English abstract).
- Chung, C.H., and Choi, D.K., (1993), Paleoclimatic implication of palynoflora from the Yeonil Group (Miocene), Pohang area, Korea, Journal of the Paleontological Society of Korea, 9, p143-154, (in Korean with English abstract).
- Endo, S., (1938a), Cenozoic plants from Tyosen (Korea) (I), Journal of the Geological Society of Japan, 45, p85-90, (in Japanese).
- Endo, S., (1938b), Cenozoic plants from Tyosen (Korea) (II), Journal of the Geological Society of Japan, 45, p326-328, (in Japanese).
- Endo, S., (1939), Some new and interesting Miocene plants from Tyosen (Korea), Prof. Yabe‘s 60th birthday, Tokyo, p333-349, pl. 23.
- Endo, S., (1950), On the fossil Carpinus from Japan and Korea, Short Papers IGPS, (2), p51-57, pl. 6.
- Flora of North America Editorial Committee, (1997), Flora of North America North of Mexico, 3, Magnoliophyta: Magnoliidae and Hamamelidae, Oxford University Press, Oxford, p616.
- Hillier, J., and Coombes, A., (2002), The Hillier manual of tree & shrubs, 7th ed, Hillier Nurseries, Winchester, British, p512.
- Hu, H.H., and Chaney, R.W.,, (1940), A Miocene flora from Shantung Province, China. Part 1, A Miocene flora from Shantung Province, China. Part 1, 507, p1-82, pls. 1-50.
- Huzioka, K., (1943), On some fossil involucres of Ostrya and Carpinus from the Miocene deposits of Hokkaido and Tyosen, Journal of the Geological Society of Japan, 50, p285-291, (in Japanese).
- Huzioka, K., (1954), Notes on some Tertiary plants from Tyosen (Korea), III Transactions & Proceedings of Palaeontological Society of Japan, N.S.(13), p117-123, pl. 13.
- Huzioka, K., (1972), The Tertiary floras of Korea, Journal of Mining College, Akita University, Ser. A, 5, p1-83.
- Institute of Botany and Institute, Geology and Palaeontology of Academy Sinica, (1978), Cenozoic plants from China, Fossil plants of China, 3, Science Press, Beijing, p232, (in Chinese).
- Jeon, J.I., and Chang, C.S., (1997), Reconsideration of Carpinus L., (Betulaceae) of Korea primarily based on quantitative characters, Korean Journal of Plant Taxonomy, 27, p157-187, (in Korean).
- Jeon, J.I., and Chang, C.S., (2000), Foliar flavonoids of genus Carpinus in eastern Asia - Primarily based on native taxa to Korea -, Korean Journal of Plant Taxonomy, 30, p139-153, (in Korean).
- Jones, S.B., and Luchsinger, A.E., (1986), Plant systematics, Mcgraw-Hill Book Company, USA, p512.
-
Jeong, E.K., Kim, K., Suzuki, M., and Kim, J.W., (2009), Fossil woods from the Lower Coal-bearing Formation of the Janggi Group (Early Miocene) in the Pohang Basin, Korea, Review of Palaeobotany and Palynology, 153, p124-138.
[https://doi.org/10.1016/j.revpalbo.2008.07.006]
- Kanehara, K., (1935), On the geological age of the Yeonil Series, Journal of the Geological Society of Japan, 42, p334-335, (in Japanese).
- Kanehara, K., (1936), On the geological study of the northern part of Yeonil district, Kyeongsangbug-do, Korea, Journal of the Geological Society of Japan, 43, p73-103, (in Japanese).
- Kim, B.K., (1987), Cenozoic Erathem, in Lee, D.S. (ed.), Geology of Korea, Geological Society of Korea, p202-221.
- Kim, J.-H., (1996), Present status and prospect of Korean Paleobotany, Journal of Geography, 22, p91-105, (in Korean with English abstract).
-
Kim, J.-H., and Choi, S.-I., (2008), Discussion on the Metasequoia fossils from the Miocene Keumkwangdong Formation of the Janggi Group, Korea, Journal of the Korean Earth Science Society, 29, p319-327, (in Korean with English abstract).
[https://doi.org/10.5467/JKESS.2008.29.4.319]
-
Kim, J.-H., Nam, K.-S., Jeon, Y.-S., (2017), Diversity of Miocene fossil Acer from the Pohang Basin, Korea, Journal of the Geological Society of Korea, 53, p387-405, (in Korean with English abstract).
[https://doi.org/10.14770/jgsk.2017.53.3.387]
- Kim, K., Jeong, E.K., Sun, B.Y., and Lee, J., (2008), New record of fossil woods from the Janggi Group in Pohang, Korea, Journal of the Paleontological Society of Korea, 24, p135-147, (in Korean with English abstract).
- Kim, W.H., (1990), Significance of Early to Middle planktonic foraminiferal biostratigraphy of the E-core in the Pohang basin, Korea, Journal of the Paleontological Society of Korea, 6, p144-164.
- Ko, K.S., (1991), Taxonomy of vascular plants, Saemunsa, Seoul, p648, (in Korean).
- Kryshtofovich, A., (1921), Contributions to the Tertiary flora of eastern Asia. 2. On the Tertiary flora of in Korea, Records of the Geological Committee of the Russiaic Far East, 18, p1-14, (in Russian).
- Lee, H.Y., (1987), Korean paleontology, Mineumsa, Seoul, Korea, p434, (in Korean).
- Lee, H.W., Jung, W.-S., Kim, H.-G., and Lee, S.-H., (2003), An analysis of climatic elements around the Pohang area, Journal of the Korean Earth Science Society, 24, p449-455, (in Korean with English abstract).
- Lee, T.B., (2006), Coloured flora of Korea, Hyangmoon Publishing Co., Seoul, p914.
- Lee, Y.D., and Lee, Y.G., (1986), A study on the Neogene Tertiary in Pohang Basin, South Korea, Journal of Science, Pusan National University, 42, p281-297, (in Korean).
- Lee, Y.S., and Lee, S.T., (1991), Modern plant taxonomy, Useong Publishing Co., Seoul, p509, (in Korean).
- Lee, H.K., Moon, H.-S., Min, K.D., Kim, I.-S., Yun, H., and Itaya, T., (1992), Paleomagnetism, stratigraphy and geologic structure of the Tertiary Pohang and Changgi Basins; K-Ar ages for the volcanic rocks, Journal of the Korean Institute of Mining Geology, 25, p337-349, (in Korean with English Abstract).
- Lee, Y.G., You, H.S., and Koh, Y.K., (1991), Biostratigraphy and paleoenvironments of Yeonil Group in Pohang area, Journal of the Paleontological Society of Korea, 7, p32-62, (in Korean with English abstract).
- Lim, K.H., Jang, C.B., Kwon, J.R., Lee, H.W., Lee, H.S., Kim, C.G., and Park, J.N., (1994), The Fossils of Chosen 3, Science Engineering Press, Pyongyang, North Korea, p223, pls. 1-96 (in Korean).
- Nathorst, A.G., (1883), Contribution à la flore fossile du Japon, Kongl Svenska Vetenskps Akademiens Handlinger, 20, p3-92.
-
Pigg, K.B., Manchester, S.R., and Wehr, W.C., (2003), Corylus, Carpinus and Palaeocarpinus (Betulaceae) from the middle Eocene Klondik Mountain and Allenbyformations of northwestern North America, International Journal of Plant Science, 164, p807-822.
[https://doi.org/10.1086/376816]
-
Stults, D.Z., Axsmith, B.J., and Haywick, D., (2002), Evidence of Carpinus (Betulaceae) in the Late Tertiary (Pliocene) of Alabama, American Journal of Botany, 89, p1547-1549.
[https://doi.org/10.3732/ajb.89.9.1547]
- Sun, B.-N., Cong, P.-Y., Yan, D.-F., and Xie, S.-P., (2003), Cuticular structure of two angiosperm fossils in Neogene from Tengchong, Yunnan Province and its paleoenvironmental significance, Acta Palaeontologica Sinica, 42, p216-222, (in Chinese).
- Takahashi, K., and Kim, B.K., (1979), Palynology of the Miocene Formations in the Yeongil Bay district, Korea, Palaeontographica, Abt. B, 170, p10-80.
- Tanai, T., (1952), Notes a propose de quelques plantes fossils dans le groupe D'Ennichi (Yongil) du Corée Méridionale. I, Transactions & Proceedings of Palaeontological Society of Japan, N.S.(8), p231-236, pl. 22.
- Tanai, T., (1955), Illustrated catalogue of Tertiary plants in Japanese Coal Fields. 1. Early and Middle Miocene floras, Geological Survey of Japan, Report, 163, p1-61, pls. 1-22 (in Japanese with English summary).
- Tanai, T., (1961), Neogene floral change in Japan, Journal of Faculty of Science, Hokkaido University, Ser, 4, 11, p119-398, pls. 1-32.
- Tanai, T., (1964), Neogene floral change in Japan, 10th International Botanical Congress, Edinburgh, p1-6.
- Tanai, T., (1972), Tertiary history of vegetations in Japan, In A. Graham (ed.), Vegetation and vegetational history of northern Latin America, Elsevier, Amsterdam, The Netherlands, p235-255.
- Tanai, T., (1984), Recent progress of angiosperm paleobotany, Journal of Earth Science, 93, p76-84, (in Japanese).
- Tanai, T., (1991), Tertiary climate and vegetational changes in the Northern Hemisphere, Journal of Geography, 100(6), p951-966.
- Tanai, T., and Onoe, T., (1959), A Miocene flora from the northern part of the Joban Coal-Field, Japan, Bulletin of Geological Survey of Japan, 10, p261-286, pls. 1-7 (in Japanese).
- Tanai, T., and Onoe, T., (1961), Mio-Pliocene flora from the Ningyo-toge area on the border between Tottori and Okayama Prefectures, Japan, Report of Geological Survey of Japan, (187), p1-62, pls. 1-18.
- Tateiwa, I., (1924a), Tertiary plants from Eunnichi and Choki, N. Keisho-do, Journal of Chosen National History and Society, Reading paper, (1), p36-57, (in Japanese).
- Tateiwa, I., (1924b), Geological atlas of Chosen, no. 2, Eunnichi, Kyuryuho and Choyo sheets (1:50,000), Geological Survey of Chosen, p6, 3 maps (in Japanese).
- Tateiwa, I., (1976), The Korea-Japanese geotectonic zone. New interpretations on the geotectonic development of the Far East continentalterritories and the insular arcs of Japan, with special reference to the history of geological research in Korea, University of Tokyo Press, Japan, p654, (in Japanese).
- Uemura, K., and Tanai, K., (1993), Betulaceae leaves and fruits from the Oligocene of Kitami, Hokkaido, Japan, Memories of the National Science Museum, Tokyo, 26, p21-29.
- Um, S.H., Lee, D.W., and Bak, B.S., (1964), Explanatory text of the geological map of Pohang sheet, Geological Survey of Korea, Seoul, Korea, p17, (in Korean with English abstract).
-
Wilde, V., and Frankenhäuser, (1998), The Middle Eocene plants taphocoenosis from Eckfeld (Eifel, Germany), Review of Palaeobotany and Palynology, 161, p7-28.
[https://doi.org/10.1016/S0034-6667(97)00067-5]
- Wolfe, J.A., (1978), A paleobotanical interpretation of Tertiary climates in the northern Hemisphere, American Scientist, 66, p694-703.
-
Worobiec, G., and Szynkiewicz, A., (2007), Betulaceae leaves in Miocene deposits of the Beƚchatόw Lignite Mine (Central Poland), Review of Palaeobotany & Palynology, 147, p28-59.
[https://doi.org/10.1016/j.revpalbo.2007.06.001]
- Yamanoi, T., (1992), The palyno-flora of Early Middle Miocene sediments in the Pohang and Yangnam Basins, Korea, In Ishizaki, K., and Saito, T. (eds.), Centenary of Japanese Micropaleontology, Terra Scientific Publishing Company, Tokyo, Japan, p473-480.
- Yoon, S., (1975), Geology and Palaeontology of the Tertiary Pohang District, Korea, part 1. Geology, Journal of the Geological Society of Korea, 11, p187-214 .
- Yoon, S., (1982), Tertiary stratigraphy of the Eoil Basin, Journal of the Geological Society of Korea, 18, p173-180.
- Yun, H., (1986), Emended stratigraphy of the Miocene Formation in the Pohang Basin, part 1, Journal of Palaeontological Society of Korea, 2, p54-69.
- Yun, H., Min, K.D., Moon, H.K., and Yi, S.S., (1991), Biostratigraphic, and tephrochronological study for the correlation of Tertiary formations in southern part of Korea: Regional tectonics and its stratigraphical implication in the Pohang Basin, Journal of Palaeontological Society of Korea, 7, p1-12.
- Yun, J.B., (2017), A pictures book of Tree APG, Jisun Publishers, Seoul, Korea, p751, (in Korean).